研究紹介

カイコとシルクの塩見研で研究している内容の一部をご紹介します。
カイコの卵休眠誘導・昆虫の季節適応に関する研究
1. カイコの卵休眠誘導について
野外の昆虫は季節の移り変わりを敏感に察知し,冬のような生存に厳しい季節や春や夏のように成長・繁殖に都合の良い季節の到来を予知し,自らだけでなく次世代個体の適応度を上げる季節適応を行なう。
休眠はその極端な例であり休眠するか否かにより一年に何世代を繰り返すかという化性が決まる。一年に2世代のライフサイクルを繰り返す昆虫は2化性と呼ばれるが,家畜化された昆虫のカイコ (Bombyx mori) にも2化性の系統が存在し,これらは母蛾が受けた環境条件により次世代卵の休眠性が決定する。
例えば,母蛾(メス成虫)の胚期を25℃・全暗(25DD)に保護すれば胚発育の初期で通常の胚発生を停止し5℃程度に一定期間曝されない限り休眠は覚醒しない。一方,15℃・全暗(15DD)に保護すれば次世代卵は非休眠卵となり約1週間で幼虫が孵化する(図.1.1,1.2)。
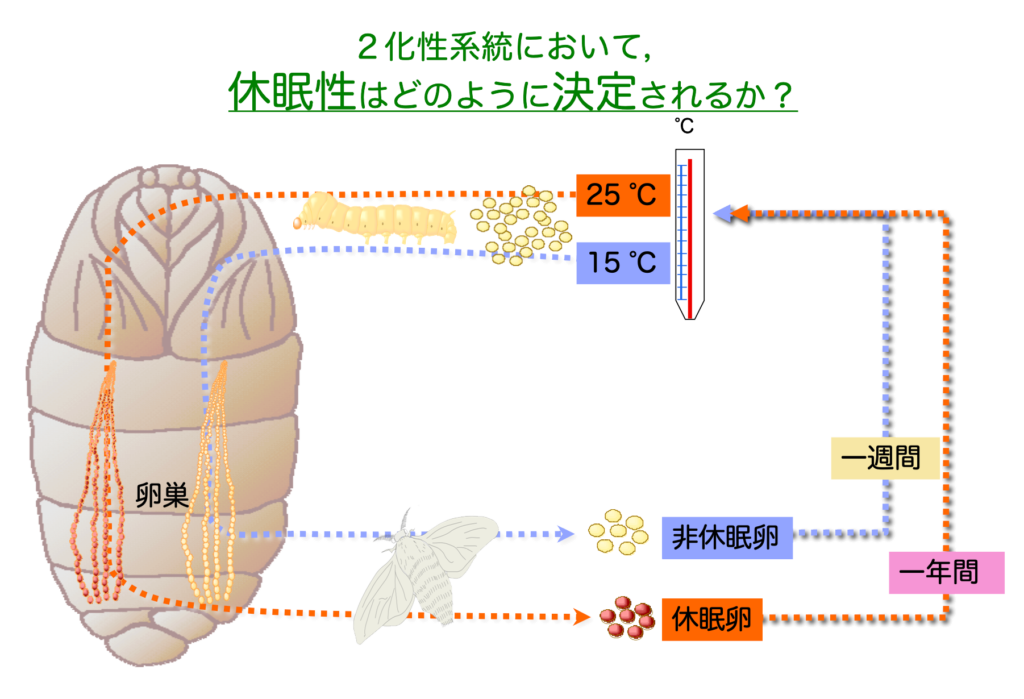
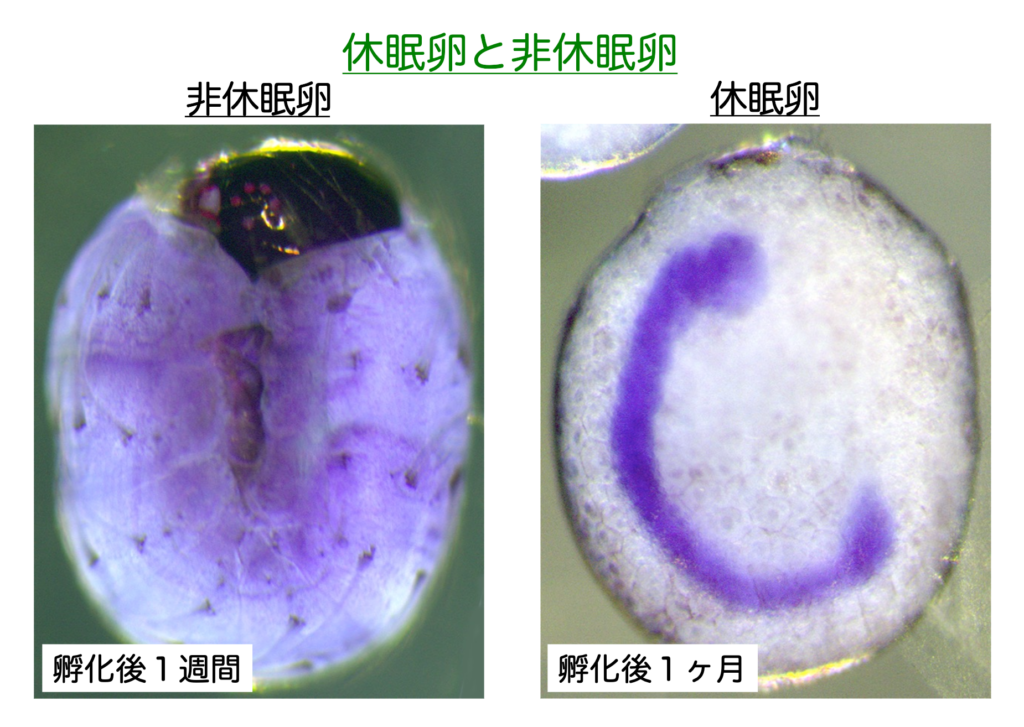
このように,カイコの胚休眠誘導は環境温の受容から現象の発現までに1ヶ月以上を有する親子2世代に渡ったユニークな環境応答機構をもつ。
このような現象は,「predictive adaptive response」(予知的適応応答)と言われ,環境刺激に対して即時に反応するのではなく環境情報の受容から現象の発現までに比較的長い期間を有する反応であり生物学的に非常に興味深い現象である。
しかし,ヒトを含めてその詳細な分子機構についてはまだ不明な点が多く,そのコントロールによる医療・産業利用が期待されている。
一方,カイコの胚休眠誘導の分子機構に関する研究については,先人たちによる偉大な報告が数多くある。
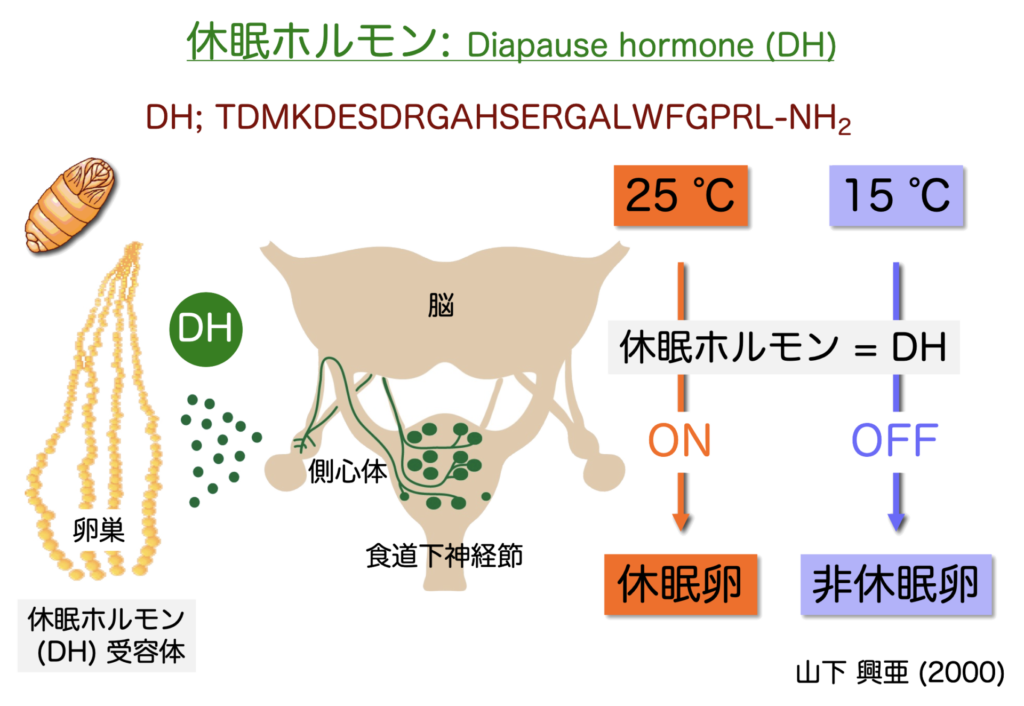
その内,「休眠ホルモン(DH)」に関する研究は特筆すべきものである。
1952年には,蛹の食道下神経節 (SG) 中に休眠卵を誘導する物質 (DH) が存在することが報告され,1991年にはDHが24残基のアミノ酸からなる神経ペプチドホルモンであることが明らかになった。
また,卵巣には DH受容体 (DHR) が存在し,血液中に放出されたDHがDHRに作用し,休眠卵が誘導されることが示された(図.1.3)。
2000年に山下 興亜先生らは,これまでの休眠研究を「休眠(ホルモン)物語」としてまとめ,カイコのユニークなライフサイクルとその分子基盤を俯瞰した。
2. 環境温の受容から DH 放出までの1ヶ月の道のり
温度センサーであるイオンチャネルの TRPA1は,数種の昆虫において侵害刺激となる高温に対する即時的な警報(アラート)の役割を担っている。
一方,カイコのオーソログ(BmTRPA1)は,20℃以上で活性化し,DH 放出の制御を介して次世代卵の休眠性を決定する。
つまり,母蛾の胚期の温度感受性期にBmTRPA1が25°Cと15°Cの温度に違いを感知し,25°Cの場合,このセンサー分子の活性化が誘起され,やがて蛹期に DH シグナル経路の活性化が起こり休眠が誘導される。
次に蛹期においては,脳の特定の領域からの神経支配により,25DDではDHの血液中への放出の促進が,15DDではその抑制が起こり,休眠性が決定されると1950年代から考えられていた。2021年には,蛹の脳において,DH 放出の上位で脳内の神経ペプチドであるコラゾニン(Crz)が働き,DHの放出促進に関与していることがわかった(図.1.4)。
また,Crzの分泌はGABA性神経伝達により抑制されていることも示した。
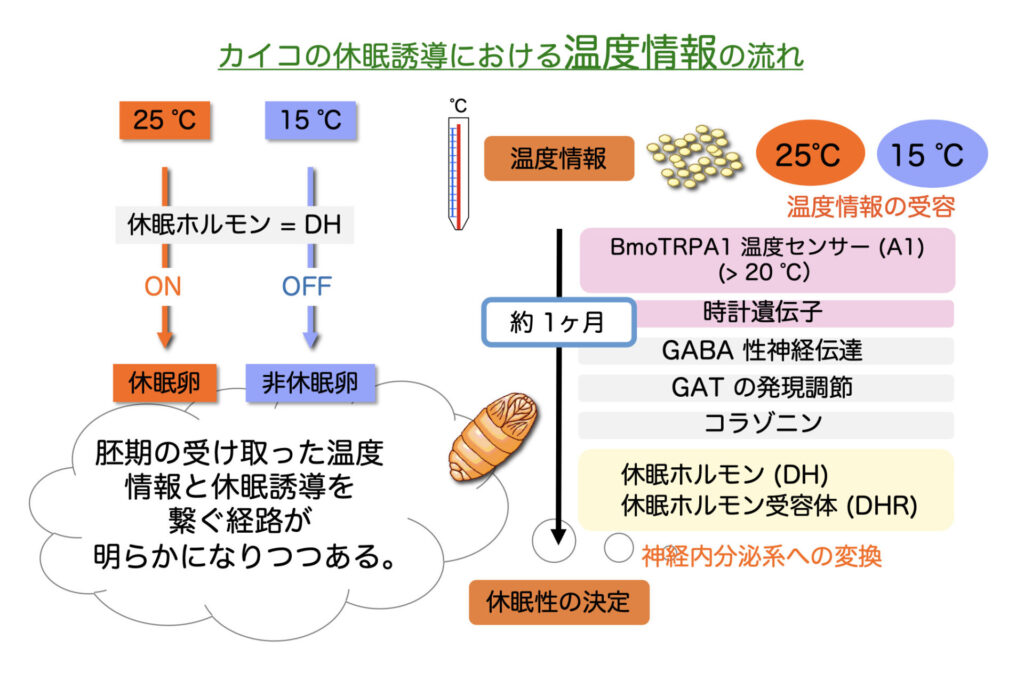
さらに,蛹期の脳-SG複合体における細胞性GABAトランスポーター (GAT1)の遺伝子発現量は,15DDでは25DDに比べて極端に低いことがわかった。
GAT1は,シナプス間隙のGABAを回収する働きを持つため,15DDではGABAの回収が行われず,GABA性神経伝達が持続的に行われ,非休眠卵の産生に繋がっていると考えられた。
さらに,2022年にはGABA性神経伝達の上位で時計遺伝子による調整が行われることを明らかにした。
さらに胚期から蛹期に至る約1ヶ月間に環境温の情報がどのように「保存・伝達」に関わりが予想される候補遺伝子群の KO 系統を作出しており,休眠性に影響のある遺伝子を見つけている。
これらの研究をもとに環境温の受容からDH放出に繋がる分子機構の全貌を今後明らかにしたい。
3. 昆虫の季節適応の分子機構の解析
昆虫では,特にチョウの成虫の翅パターンの季節型は多くの人を魅了し,よく知られている。これらの季節的差異は,成虫の行動や生理状態と関係しており,体温調節や対捕食者戦略(隠蔽色や目玉模様など)として機能していることが示されているが,その適応的意義は不明な点も多い。また,カイコで示したように多くの昆虫は休眠とよばれる通常とはまったく異なったライフスタイルを冬が訪れる前に整える。
リンゴの樹の害虫として知られるヒメシロモンドクガ (Orgyia thyellina) は,幼虫期の日長条件の違いにより,終齢幼虫や蛹の体色,繭色,翅サイズ,卵の休眠性,卵径,卵色,卵殻の厚さ,営繭行動などにさまざまな季節的多型を示す(図.1.5)。

季節的多型を示す昆虫は数多く知られているがこれほどまでにさまざまな表現型多型を現わす種は珍しい。長日条件では,幼虫や蛹は淡い体色を呈し,薄白色の繭を葉の裏に作る。さらに雌の成虫は長翅型となり,白黄色で卵殻が薄く,卵径の小さな非休眠卵を多数産生する。一方,短日条件では幼虫や蛹の体色は濃くなり,黒褐色の繭を木の幹などの風下に作る。そして雌の成虫は短翅型となり,飛翔筋が長翅型の雌に比べて著しく矮小化し,飛翔能力を持たない。また,卵は長翅型の産生する卵に比べて色が濃く,卵殻も厚く,卵径が大きく,卵重も重い休眠卵を少数産生する。これらの表現型は雌のみで誘導され,雄では観察されない。雄の翅は日長条件が変化しても常に長翅型であり,性的二型を示す。このようなさまざまな季節的多型の発現には,このドクガが日長の変化を感じ取り,その後に訪れる季節を予知して,一生を通じてその時期に最適な生存戦略を行っているからであると推測される。つまり,長日条件であり生存に適した初夏の時期には,熱吸収の抑制や,生存範囲の拡大,次世代の個体数の増加などを行ない,短日条件である秋口には,その後訪れる生存に不適切な冬に備え,熱吸収の促進や,落葉や風害への対策,越冬するために必要な耐凍性や休眠性の獲得など行なっていると推測される。
カイコと同様に胚発生の初期に休眠を開始する昆虫であり,DH によって胚休眠が誘導される。カイコの胚休眠誘導と類似した分子ネットワークにより休眠が誘導されると推測されるため,カイコで明らかになった知見を応用し,害虫防除に役立てられないか実験圃場のリンゴ園で検討している。さらに,さまざまな昆虫で幼虫や蛹休眠の分子機構の解析も進めている。これらの知見をもとに,害虫のライフサイクルを自在にコントロールして害虫防除やカイコの効率的な保存が実現できればと研究を進めている。

祖先種クワコからカイコへの進化(家畜化)に関する研究

カイコは祖先種のクワコ (Bombyx mandarina) から 5千 ~ 1万年前に家畜化したとされる。家畜化に伴い絹糸生産のための有用形質が人為選抜されており,繭量の増加に加えて集約的な飼育,発育タイミングの斉一化などクワコおよび野外の昆虫では観察されない家畜化形質を保有している。
その一方でカイコはクワコがもつ季節応答機構を喪失していることがある。カイコとクワコは遺伝子の並び方の相同性(シンテニー)が高度に保存されており交配・継代が可能である。
そのためクワコからカイコの家畜化の過程で生じた僅かな遺伝的変異で野生種から家畜種への劇的な差異が生じたと考えられる。その一例が休眠の誘導である。
クワコもカイコと同様に胚休眠を行なうが,休眠性の決定には母蛾の胚期の温度条件は関係せず幼虫期の日長条件が重要となる。
この違いはカイコが家畜化の過程において新たな環境応答機構を獲得した結果であると推測している。
日本の本州に生息するクワコは休眠卵と非休眠卵を産み分けており,桑のある時期は非休眠卵を,冬が来る前には休眠卵を産卵して冬は休眠卵で越冬する。
クワコの休眠性は母蛾が幼虫期に受けた光周期(日長)によって決定し, 16時間明条件:8時間暗条件 (16L8D) のような長日条件では非休眠卵となり,8L16D のような短日条件では休眠卵が産まれる。
クワコにもカイコと同じ様な DH と DHR があり,これらによる DH シグナル経路により休眠卵が誘導されることを明らかにし,クワコの母蛾にもカイコと同じ様な温度センサー (BmaTRPA1) が存在し,25℃で活性化されることを明らかにしている。つまり,カイコとクワコでは休眠誘導のキッカケとなる環境情報は異なるわけだが,その仕組みはよく似ているように思える。それではカイコとクワコで休眠を誘導するしくみはどのように異なっているのだろうか?このことを調べるためにゲノム編集技術を利用し,カイコの温度センサーの KO 系統を作出した。この系統では休眠誘導に関わる胚期での温度受容ができないと考えられるが,その休眠性はクワコと同じ様に幼虫の日長条件により休眠性が決定されることがわかった(図.2.1)。
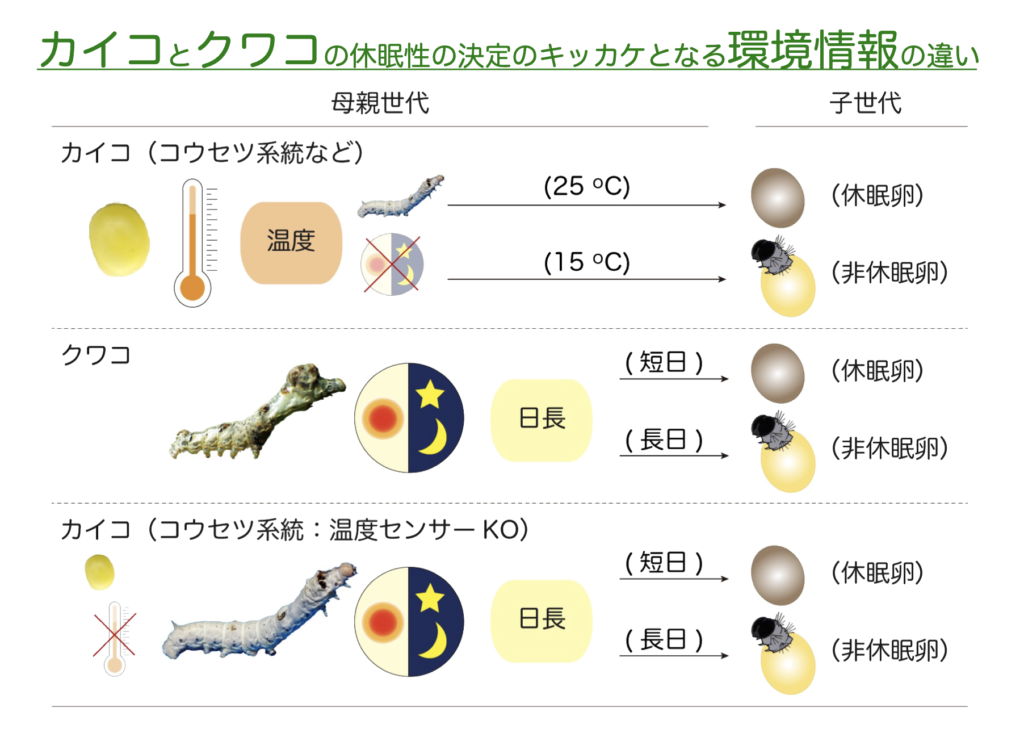
つまり,短日条件で休眠卵,長日条件で非休眠卵が産卵された。おそらく,カイコ(コウセツ系統)の野生型では温度情報および BmoTRPA1 の活性化シグナルと休眠誘導に関わる何らかの装置に強い繋がりがあるが,KO 系統では,この繋がりが解除され,カイコの祖先種のクワコと同じように幼虫期の光周期依存的な休眠性の決定機構が働いたと推測できる。
また,コウセツ系統のような温度に依存して休眠性を決定する系統は,クワコからカイコが家畜化される過程において,人為的に日長管理よりも温度管理が容易なことから家畜化における人為選抜の過程でカイコ集団内に広がったのではないかと推測している。
つまり,休眠に関しても野外の昆虫とは異なる意義をもつ。野外(温帯域)の昆虫の休眠の意義の一つは越冬により,生存に不適な環境へ適応することであるが,カイコにおいては,越冬というよりもむしろ,産業活動における蚕種(カイコ卵)の効率的・計画的な保存のためであると考えられる。
言い換えれば,カイコは卵で休眠するが,卵の時期が人類にとって最も管理しやすく,蚕種として売買の対象となりうることから,胚休眠の形質が選抜・保存され,人類が光周期よりも温度管理を行なうことが容易だったため,温度条件で優先的に休眠が誘導される形質が選抜・保存されるようになったのではないかと推測している。

休眠誘導以外にも形態・行動など色々な面でカイコとクワコでは異なる。周知の通り,カイコは人類の絹糸生産の歴史とともに継代され,飼育と絹糸生産に都合の良い形質を保つように選抜育種され家畜化(産業化)された昆虫種であり,決して自然下の野外では生存できない。当然,カイコの飼育に季節変化は関係なく,重要なのは人の手による蚕室の環境管理である。いわば,カイコが受容する最大の環境要因は人為管理・選抜と解釈でき,これによりカイコは独自の発展(進化)を遂げたと考えられる。カイコのように昆虫を産業化する際,カイコは独自の環境応答機構を獲得したのではないかと推測している。我々はこのような環境応答機構を「孫悟空が觔斗雲に乗ってどこまで飛んでもお釈迦様の掌の中だった」という中国の古典文学「西遊記」になぞらえて,「in hand な環境応答システム」(図.2.2)と名付け,その進化機構の研究を進めている。

信大シルクプラットフォームの構築
「信大シルク」プラットフォームのプロジェクトがいよいよ始動しました。
